
Diatomáceas – as algas douradas
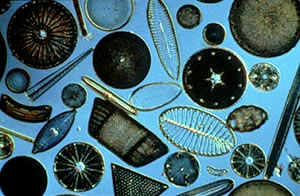
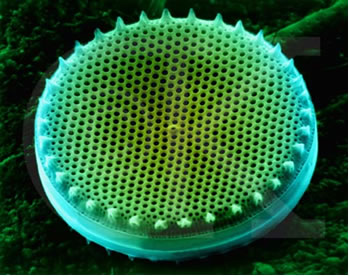
Diatomáceas são microrganismos eucariontes unicelulares que ocorrem nos mais diversos ambientes úmidos e aquáticos, suspensos na coluna d´água ou aderidos a diversos substratos: macrófitas (epifíticas), rochas (epilíticas), animais (epizóicas), grãos de areia (episâmicas), sedimento (epipélicas). São fotossintetizantes, possuindo clorofilas do tipo a e c, mas algumas poucas espécies são capazes de resistir heterotroficamente a condições de pouca luz e de baixa disponibilidade de matéria orgânica(Fig. 1).

Figura 1. Valvas de diatomáceas marinhas em lâmina confeccionada pelo espanhol Muñoz Moraton, coleção do Prof. Hermes Moreira-Filho .Foto de Kaoli Cavalcante
A principal característica morfológica das diatomáceas é a parede celular impregnada de sílica (SIO2.nH2O), envolvida por uma fina camada de matéria orgânica, conhecida como FRÚSTULA. É altamente diferenciada, ornamentada por diferentes tipos de estruturas, e sempre dividida em duas unidades chamadas TECAS, as quais se encaixam como duas placas de petri (Fig. 2). A teca maior é conhecida como EPITECA (sempre originada da célula mãe), enquanto a menor é chamada HIPOTECA. Cada uma das tecas é composta pela VALVA e pelo CÍNGULO. As valvas são localizadas nas extremidades da célula e se unem por várias unidades silíceas delicadas, as bandas do cíngulo. Cada valva é composta por superfície e manto. Uma variedade de estruturas e projeções ornamentam as frústulas, as quais são a base da taxonomia das diatomáceas. Para conhecer as principais estruturas das frústulas da diatomácea, clique AQUI.

Figura 2. Estrutura da frústula de diatomáceas. Ilustração de Kaoli Cavalcante
Várias diatomáceas secretam polissacarídeos através de estruturas especializadas da valva (fultopórtulas, rimopórtulas, ocelos etc.), possibilitando às células a formação de cadeias, adesão a substratos e motilidade sobre superfícies. Tais secreções, quando hidratadas no meio, podem ocorrer em forma de tubos de mucilagem (Fig. 3), almofadas (Fig. 4, 5), ou pedúnculos de mucilagem.


Figura 3. Diatomáceas formando tubo de mucilagem Foto de Mariana Feres.
Figura 4. Diatomáceas aderidas a substrato por almofadas. Foto de Eloési dos Santos



Figura 5. Diatomáceas epífitas sobre outras diatomáceas (Hydrosera) aderidas por almofadas de mucilagem. Foto de Cleto Peres
Figura 6. Diatomáceas (Gomphonema) formando pedúnculos de mucilagem. Foto de Eloési dos Santos
Figura 7. Diatomáceas epífitas vistas em MEV. Observe pedúnculos de mucilagem formados por Gomphonema. Foto de Thelma Ludwig.
Mucilagem pode ser observada em microscopia óptica (Fig. 3, 6) e eletrônica (Fig. 5, 7) com técnicas de preparação para amostras não oxidadas, enquanto que a morfologia e ultraestrutura dos componentes da frústula são melhor observáveis em material limpo, através da oxidação da matéria orgânica (Fig. 8). Técnicas de oxidação da matéria orgânica podem ser acessadas neste link.
As diatomáceas também possuem uma forma peculiar de reprodução assexuada, que conduz à redução gradativa da dimensão celular. Para restaurar o tamanho original da espécie, ocorre a formação de células zigóticas especiais, durante a reprodução sexuada. O ciclo de vida é do tipo haplobionte diplonte, no qual os indivíduos adultos são diplóides (2n) ve a meiose ocorre para formação dos gametas (meiose gamética; aprenda sobre tipos de ciclos de vida nas algas).


Figura 8. Frústula de Cymbella aspera observada em material fresco (esquerda) e valva depois de oxidada (direita), com melhor observação de suas ornamentações. Fotos de Thelma Ludwig (esquerda) e Priscila Tremarin (direita)
Na reprodução assexuada, as células adultas (2n) dividem-se, formando duas células-filhas. Cada uma destas herda da célula-mãe uma teca e a outra teca é formada durante o processo de divisão celular. A teca herdada da célula-mãe sempre será a EPITECA, enquanto a teca formada no processo de divisão, a HIPOTECA, será menor. Consequentemente, a célula-filha que herdou a epiteca da célula-mãe possuirá o mesmo tamanho da original, mas a outra sofrerá redução de tamanho, em relação à primeira (Fig. 9).

Figura 9. Modelo esquemático de reprodução assexuada em diatomáceas. Ilustração de Kaoli Cavalcante

Figura 10. Variação populacional em Nitzschia frustulum. Fotos de Kaoli Cavalcante
Após sucessivas gerações, o resultado deste processo será a redução do tamanho das células de uma parte da população (aquelas que herdaram as hipovalvas das células-mães, Fig. 10). Quando atingido um limite, diferente para cada espécie, as diatomáceas entram em reprodução sexuada , restabelecendo o tamanho original das células, da seguinte forma: células adultas (2n) produzem gametas (n) por meio de meiose . Após a fusão de gametas (plasmogamia, mas sem a fusão de núcleos), uma célula desenvolve-se e se expande em volume – o auxósporo – e então, ocorre a fusão dos núcleos (cariogamia) e a formação do zigoto (2n), antes da formação da nova frústula. É via auxósporo que o tamanho original da espécie é restabelecido, após as sucessivas divisões assexuadas.
O auxósporo não é um esporo, nem um estágio de resistência. Confusão também é feita entre os termos zigoto, auxósporo e célula inicial.
Auxósporo, segundo Round et al. (1990), é a célula binucleada formada a partir da ocorrência de plasmogamia (ou do final da meiose, em espécies autógamas), até o momento em que ocorre a fusão de núcleos (Fig. 11, 12).


Figura 11. Auxósporo de Thalassiosira rudis. Foto de Cristiane Szawka.
Figura. 12. Auxósporo de Aulacoseira ambigua. Foto de Priscila Tremarin
O zigoto seria, portanto, a fase seguinte à cariogamia.
Células iniciais aparecem a partir do desenvolvimento do zigoto, quando a parede celular do auxósporo (uma parede orgânica formada de polissacarídeos) gradualmente é impregnada de sílica. A parede celular formada neste estágio (perizônio) é morfologicamente diferente da parede celular de células vegetativas (Fig. 13).


Figura 13. Células iniciais de Aulacoseira ambigua. Foto de Priscila Tremarin.
No protoplasto, as organelas mais conspícuas são os plastos, normalmente marrom-dourados, devido à coloração de pigmentos acessórios, que mascaram a cor verde das clorofilas a e c. Estes pigmentos são os carotenóides, principalmente β-caroteno, diatoxantina, diadinoxantina e fucoxantina. A forma, número e posição dos plastos podem auxiliar na taxonomia do grupo (Fig. 14).





Figura 14. Exemplos de diferentes tipos de plastos em diatomáceas. Da esquerda para a direita: Melosira varians, Aulacoseira granulata, Asterionella formosa, Cymbella aspera (fotos de Thelma Ludwig) e Navicula sp. (foto de Kaoli Cavalcante).
As mitocôndrias e outras organelas posicionam-se geralmente na região periférica do protoplasto, entre a parede celular e um ou dois grandes vacúolos centrais. O núcleo (Fig. 15) geralmente é bem visível, central em algumas células e parietal em outras.

Figura 15. Thalassiosira rudis com núcleo evidente. Foto de Cristiane Szawka.
Além destas, corpúsculos de óleo (Fig. 16) e reserva de polissacarídeos são frequentemente visíveis no protoplasma de células vivas. O polissacarídeo de reserva das diatomáceas é a crisolaminarina.

Figura 16. Cymbella aspera apresentando corpúsculos de óleo nas extremidades da célula. Foto de Thelma Ludwig.
Motilidade
A motilidade das células das diatomáceas ocorre particularmente nos gametas uniflagelados das cêntricas (únicas células móveis de todo o grupo!) e nas formas penadas que possuem sistema de rafe LINK PARA GLOSSÁRIO. Entretanto, há registros de movimento em algumas diatomáceas arrafídeas e de movimento rotacional nas cêntricas.O movimento de deslizamento realizado pelas diatomáceas rafídeas é bem estudado e explicado pela secreção de material mucilaginoso através da rafe (Fig. 17). O movimento é direcional, seguindo o mesmo plano da rafe – movimentos retos, curvados ou sigmóides são relativos às rafes retas, curvadas ou sigmóides, respectivamente – com alternantes acelerações, podendo ocorrer movimento de volta. É importante entender que a motilidade das células só ocorre sobre um substrato sólido. O mecanismo intracelular que origina o movimento é detalhado a seguir.
Basicamente, mucopolissacarídeos são secretados no interior da célula e atravessam a membrana celular, alcançando o meio externo via fenda da rafe. No interior da célula, filamentos de actina (proteína contrátil) estabelecem ligação com as proteínas transmembrana, que por sua vez ligam-se às moléculas de mucopolissacarídeo. No contato com o meio, estas estruturas hidratam e se fixam ao substrato. Elas estão, portanto, livres no interior da célula e na fissura da rafe, mas fixadas ao substrato. São os movimentos dos filamentos de actina presentes na periferia da célula, movimentando o conteúdo citoplasmático, em interação com as estruturas protêicas transmembranas, que fazem com que a diatomácea movimente-se em relação ao substrato. Na região distal da rafe, a mucilagem destaca-se da célula na helictoglossa, deixando um rastro mucoso e efêmero.

Figura 17. Esquema de mecanismo de motilidade nas diatomáceas com rafe. Ilustração de Kaoli Cavalcante
Motilidade é uma estratégia ecológica importante para as diatomáceas perifíticas que, com limitada dispersão, podem buscar microhabitats com melhores condições para o crescimento, especialmente de luz e de nutrientes.